
Chapter 9 ~ Ecology: From Individuals to the Biosphere
Key Concepts
After completing this chapter, you will be able to
- Describe how species are adapted to different levels of stress and disturbance in their habitat.
- Explain how population growth occurs, as well as constraints on population size.
- List major environmental factors that influence ecological communities.
- Describe what is meant by a landscape (and seascape) and how environmental influences affect their spatial and temporal dynamics.
- Outline the Gaia hypothesis and discuss its applicability to the functioning of the biosphere.
Introduction
Ecology is the study of the relationships between organisms and their environment. In the sense meant here, “environment” includes both (a) non-living factors, such as temperature, moisture, nutrients, and physical disturbances, as well as (b) living organisms, which exert influences through competition, herbivory, predation, and disease, and by providing elements of habitat (as when trees provide habitat for species living in a forest). Because all organisms and ecosystems are subjected to a multiplicity of influences, it can be difficult to predict the ecological effects of changes in environmental conditions.
Some environmental influences are resources that organisms can exploit as opportunities, which allows them to gain the necessities of life and livelihood. Other environmental influences are stressors, or constraints on productivity and reproductive success. Many stressors operate in a continuous (chronic) fashion, as is often the case for climatic factors, soil and water pollution, and many biological interactions. Other stressors affect organisms and ecosystems as events of disturbance, which cause severe damage in a short period of time. A disturbance is followed by an extended period of ecological recovery called succession. Disturbance may be caused by natural forces such as a wildfire or windstorm, or by anthropogenic influences such as the clear-cutting of a forest or ploughing of a field.
Image 9.1. An individual organism is genetically unique and is different from other individuals of its species. This green heron (Butorides virescens) was photographed in southern Florida, but it also occurs in southern Canada. Source: B. Freedman.
Image 9.2. This population of northern gannets (Morus bassanus) breeds at Cape St. Mary’s in Newfoundland. Source: B. Freedman.


Ecology considers the structure and function of the web of life at a hierarchy of levels:
- An individual organism is defined, in an evolutionary context, as a genetically unique entity. However, some species propagate by asexual mechanisms, and they may develop clones of genetically identical “individuals”.
- A population is a group of individuals of the same species that are co-occurring in time and space and can potentially interbreed with each other.
- A species consists of one or more populations in which individuals can potentially interbreed, and are reproductively isolated from other such groups.
- A community is an assembly of populations of various species that co-exist and interact as a distinctive grouping.
- An ecoscape is a spatial integration of various kinds of communities over a large area. Each community is a spatial “patch” and the ecsoscape comprises a dynamic mosaic, which is referred to as a landscape in terrestrial environments and as a seascape in marine ones.
- The biosphere consists of all of life and ecosystems on Earth and the environments where they occur.
Each of these levels of ecology is meaningful, and all are relevant to environmental science. However, these various tiers of ecology are not totally discrete—they are all interconnected and each level influences every other. This chapter examines issues that are relevant to the various hierarchical levels of ecology.
Image 9.3. Coral reefs are shallow-water ecosystems in tropical seas, and they are extremely rich in species, as is illustrated by this community near Puerto Morelos, Mexico. Source: A. Pinder

Individuals and Species
Autecology is the field within ecology that deals with the study of individuals, populations, and species. Important topics in autecology include the following:
- differences among species in life-history characteristics and in adaptations to various kinds of environmental conditions
- influences of the environment on individual organisms, including effects on their development and behaviour
- the causes of changes in the size and makeup of populations
Life-History Characteristics
Each species is unique and can be described by its anatomical, behavioural, biochemical, and ecological attributes. These characteristics are ultimately determined by the collective genetic variation that exists among the individuals that comprise the species.
Each species is unique. Nevertheless, species can be aggregated into groups based on similarities of their attributes. These affinities may be due to ancestral relatedness, due to related species sharing aspects of their evolutionary history. For example, all maple trees (genus Acer) look rather alike and occur in habitats of temperate forest. Similarly, all members of the cat family (Felidae) bear a certain resemblance and are ecologically comparable in that all are predators, although of different prey and in different kinds of habitat. However, unrelated species may also display similar attributes, usually because they have had a history of analogous changes through a phenomenon known as evolutionary convergence (or parallel evolution). Convergence suggests that, through natural selection, unrelated species living in comparable environments may evolve to resemble each other and to play similar functional roles in their ecosystem.
There are many examples of evolutionary convergence among unrelated groups of organisms. For instance, all perennial (long-lived) plants growing in arid environments have a need to conserve moisture. This critical function is enhanced by a growth form that includes adaptations to reduce water loss, such as a cylindrical trunk and branches, tissues protected by a waxy cuticle, and no leaves. Thorniness is another useful trait in an arid environment because spines deter herbivores from consuming biomass and stores of water. Many desert-inhabiting plants have developed one or more of these adaptations, including species of cacti (family Cactaceae), euphorbs (Euphorbiaceae), and succulents (Crassulaceae). Although species in these families are not closely related in an evolutionary sense, they may resemble each other because of evolutionary convergence.
There are also examples of convergence among species of animal. One is the similarities of the timber wolf (Canis lupus) of Eurasia and North America and the marsupial wolf (thylacine, Thylacinus cynocephalus) of Australia. Another example is the groundhog (Marmota monax) of North America and the marsupial wombat (Vombatus ursinus) of Australia. Also, the penguins (family Spheniscidae) of the Southern Hemisphere are similar to the guillemots, murres, puffins, and related auks (family Alcidae) of the Northern Hemisphere.
Ecologists often categorize plant species on the basis of their autecology. One system is based on the adaptations of plants for coping with certain kinds of habitat conditions. The ecologist Philip Grime has suggested that plant strategies can be divided into three basic categories, which are determined by life history and its relationship to habitat. This system proposes that two groups of environmental factors – disturbance and stress – have a strong influence on the evolution of plant life-history strategies. Disturbance may be a frequent or uncommon occurrence, and severe or mild in its intensity. Stress is a longer-term site condition, and it can be intense if associated with an extreme shortage of moisture, light, or nutrients, or innocuous if these vital factors are well available. Any particular environment can be characterized by the importance of these two groups of factors, which results in four basic kinds of habitat conditions:
- low stress and rare disturbance
- low stress but frequent disturbance
- intense stress but rare disturbance
- intense stress and frequent disturbance
However, Grime suggests that plants exhibit only three primary life-history strategies, because they cannot cope with an environment that is both stressful and frequently disturbed (4 above). The three primary life-history strategies are:
- Competitor plants are dominant in habitats in which disturbance is rare and environmental stresses are relatively unimportant. Under such conditions, competition is the major selective influence on plant evolution and on the organization of their communities. Competitive plants are effective at acquiring resources and using them to achieve a dominant position in their community by interfering with the productivity of other plants. Useful adaptations in competitors include rapid tall growth, a spreading canopy, and a widely spreading root system—these characters help to occupy space and take advantage of resources. In addition, seedlings of many competitive plants can establish themselves beneath a closed canopy.
- Ruderals occur in frequently disturbed environments with abundant resources, so stress is not great. Ruderal plants are therefore well adapted to utilizing rich but temporary habitats. They are typically short-lived and intolerant of stress and competition. Ruderals produce large numbers of seeds, which usually have mechanisms for long-distance dispersal so that newly disturbed habitats can be colonized.
- Stress-tolerators are adapted to environments that are marginal in terms of climate, moisture, or nutrient supply, but are infrequently disturbed and therefore stable. They are typical of arctic, desert, and other stressful environments, and are generally short, slow-growing, and intolerant of competition.
Another system of categorizing organisms, more commonly applied to animals, involves two groups of life-history characteristics. One consists of longer-lived organisms that produce relatively few progeny, but invest a lot of resources in each to improve their chance of survival. These are known as K-selected species. The other group, referred to as r-selected, includes short-lived species that produce large numbers of small offspring, each of which has a relatively small chance of survival, but due to the enormous numbers it is likely that some will persist. K-selected species are dominant in relatively stable, mature habitats in which competition is the controlling influence on community structure, while r-selected species occur in younger, recently disturbed habitats in which resources are freely available and rapid population growth is possible. (The source of the “K” and “r” labels comes from the logistic equation, a fundamental element of population ecology that, for simplicity, is not examined here.)
Species can also be considered in terms of other aspects of their reproductive strategy, such as how often they reproduce. Some species have only one reproductive event during their lifetime, usually dying afterward. This type of reproduction, known as semelparous, is seen in annual and biennial plants, many insects and other invertebrates, and Pacific salmon. Most semelparous species are short-lived, but some can live for many years, gradually accumulating enough energy to sustain a massive, “big-bang” reproductive effort. Semelparous reproduction is favoured in rich habitats that are frequently disturbed, and it is common among ruderal and r-selected species.
Species that reproduce a number of times during their lives are known as iteroparous. These are typically long-lived species that live in stable habitats. Iteroparous species may produce large numbers of small offspring (r-selected), or they may produce fewer, larger young, each of which receives a substantial investment of parental resources (K-selected).
Individuals and Environment
Autecology also deals with the lives of individual organisms and how they are influenced by their physical and biological environments.
As we examined in Chapter 6, all individual organisms have a fixed complement of genetic information, known as their genotype. However, the expression of genetic information (the phenotype) is influenced by environmental conditions, a phenomenon known as phenotypic plasticity. If individuals experience difficult environmental conditions, the phenotypic expression of their genetic potential may include a suboptimal growth rate and the production of few or no progeny. In contrast, other individuals that live in a more benign environment can achieve higher productivity and have many offspring. The latter, more prolific circumstance is highly desirable in terms of an individual achieving evolutionary “success.” By definition, successful individuals have managed to maximize their fitness—their genetic contribution to future generations.
The success of an individual organism is also affected by unpredictable disturbances, which may result in injury or premature death. Even if living in a benign environment, with good access to the necessities of life, an unlucky individual may just happen to be scorched by a wildfire, devoured by a predator, debilitated by a disease, or hit by a truck.
Population Ecology
The study of populations is another aspect of autecology. The abundance of all species changes over time in response to environmental factors that affect four population-related (or demographic) variables: birth rate (BR), immigration rate (IR), death rate (DR), and emigration rate (ER). The change in population size (ΔP) during a unit of time (say, a year) is described using the following equation:
ΔP = BR – DR + IR – ER
This demographic relationship is true of all species, including humans. In some cases, isolated (or closed) populations do not receive any immigration of new individuals and do not lose any to emigration. Under such conditions, ΔP is calculated as BR – DR, a value known as the natural rate of population change.
Often, ΔP is expressed as a percentage change by dividing its value by the initial population size – for instance, a population of 100 individuals that increases by 10 in one year has a 10% annual growth rate. If the percentage change in a population is constant over time, there will be an accelerating rate of increase or decrease, called exponential change.
Imagine a circumstance in which a fertile pair of individuals manages to discover a new suitable habitat—one that has not been previously occupied by their species. Under such conditions, the founder individuals will breed and the population will grow over time. Initially, resources are abundant and do not constrain growth of the population. During that period the percentage rate of increase will be constant, being limited only by how quickly progeny can be produced (the birth rate) and themselves become fertile (the maturation rate), and countered only by any deaths that might occur. This is the maximum rate of population growth, which is limited only by the biology of the species and not by competition for resources, and it is referred to as the intrinsic rate of population increase. Any population that is growing at the intrinsic rate of increase (or indeed at any fixed percentage rate) will quickly explode in abundance (see In Detail 9.1).
Eventually, however, the population will approach the carrying capacity of the available habitat, or the population that can be supported without causing resources to become limiting, or other environmental damages. At or beyond the carrying capacity, opportunities are constrained by the limited availability of resources, and so individuals in the population must compete with each other. Intense competition results in physiological stress, which may cause a decrease to occur in the birth rate and an increase in the death rate. In some cases, the rate of population increase may then decrease to zero, which occurs when the birth rate equals the death rate. This condition is referred to as zero population growth (ZPG). If ZPG is maintained, the population size will eventually level off, perhaps at a level appropriate to the carrying capacity of the habitat. A population curve of this type is referred to as “S-shaped.”
However, the earlier exponential growth may have resulted in an abundance that exceeded what the habitat could support. Such an overpopulation would degrade the environment, resulting in a decrease in its carrying capacity. If this happens, the population size will decrease through an increase in the mortality rate, or perhaps by a surge of emigration in search of new habitats. These may result in an oscillation of abundance around the carrying capacity, or in a rapid crash in the numbers of individuals in the population. Usually, a crash takes the population to a level below the carrying capacity, creating a circumstance for renewed population growth. In small habitats, however, the crash can be massive enough to extirpate a local population.
Population ecologists have developed mathematical models of population dynamics that account for the influences of such factors as the intrinsic rate of population increase, the carrying capacity of habitats, the effects of predation and disease, and even the effects of unpredictable disturbances. These models are described in introductory textbooks of ecology and are not dealt with here in any detail. For the present purposes, there are several important points to understand about population ecology:
- Populations of all species are dynamic. They change over time due to varying rates of birth, death, immigration, and emigration.
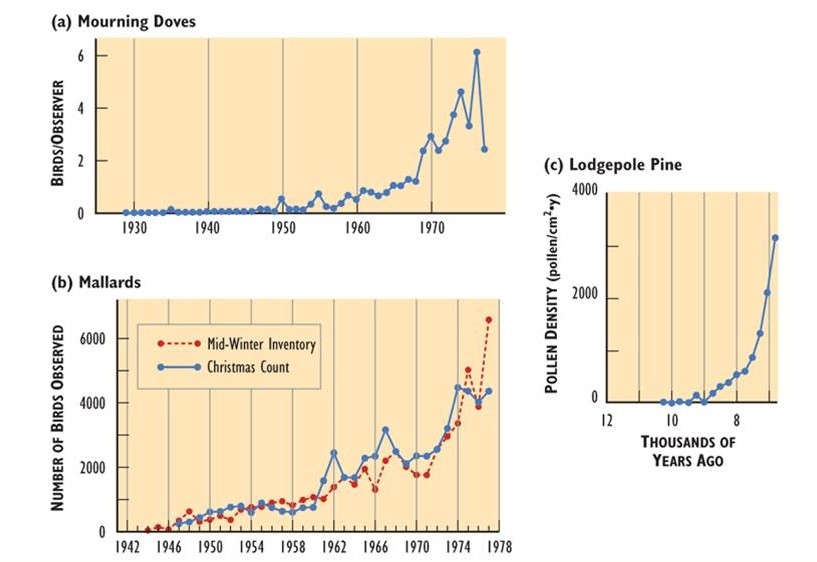
- Populations of all species can, potentially, increase rapidly under conditions in which resource availability and other factors are not constraining. Examples of rapid population growth are illustrated in Figure 9.1. However, unlimited growth cannot be sustained – in all of the cases in Figure 9.1, the population sizes eventually levelled off, decreased, or crashed.
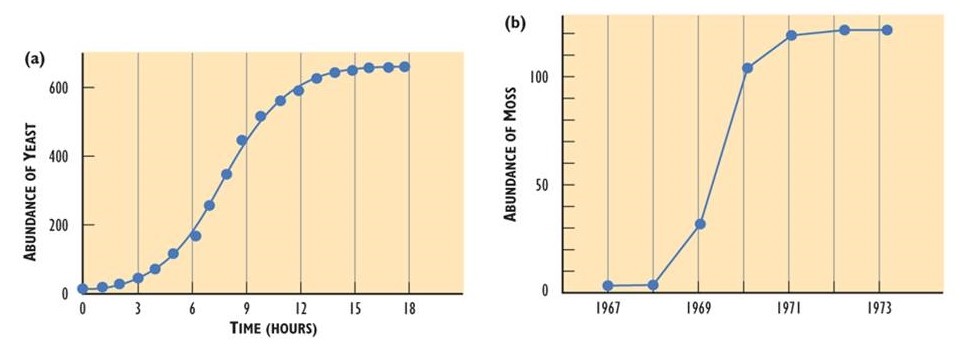
- Ultimately, the sustainable abundance of a species is limited by the carrying capacity of the available habitat. Examples of population growth that level off at the carrying capacity of the habitat are illustrated in Figure 9.2.
- Some populations are relatively stable. Usually they exist in environments in which resource availability is predictable so that a balance can be achieved with the carrying capacity. For example, relatively little change occurs in the year-to-year populations of trees growing in old-growth forest, unless a rare, catastrophic disturbance occurs.
- Other populations are relatively dynamic, changing greatly over time and rarely achieving even a short-term balance with the carrying capacity of their habitat. This is commonly true of species living in habitats that are disturbed frequently or are in an early, relatively dynamic stage of succession. Some populations are cyclic, achieving great abundances at regular intervals, interspersed by longer periods of lower abundance. Cyclic populations are obviously unstable over the short term, but they may be stable over the long term.
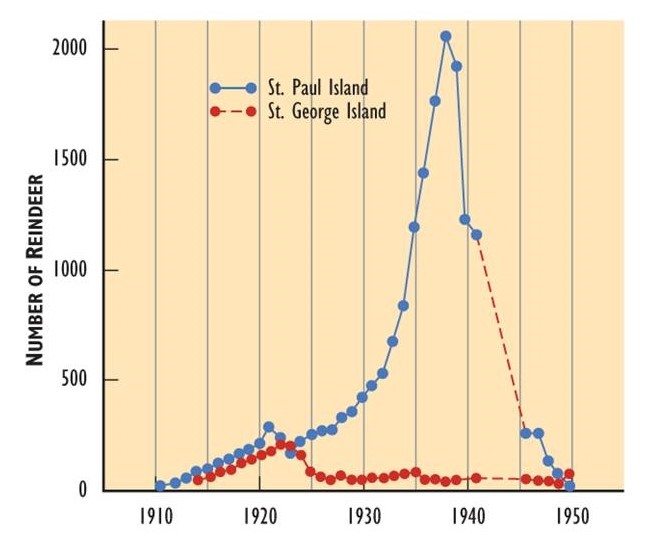
- Populations that exceed the carrying capacity of their habitat are never sustainable at that high level, because of the environmental damage is caused. Unsustainable populations eventually crash to a smaller abundance and sometimes to extinction. Figure 9.3 shows an example of rapid population growth that resulted in habitat degradation and a subsequent population crash. Populations can also crash for other reasons, such as the sudden occurrence of a deadly disease. This is happening with the native white elm (Ulmus americana) of North America, which is being decimated by an introduced pathogen (the Dutch elm disease fungus, Ceratocystis ulmi) to which this tree has little immunity. Other causes of population crashes include unsustainable levels of predation and extensive disturbances such as wildfire or clear-cutting.
Figure 9.1. Rapid Growth of Some Natural Populations. (a) The population of mourning doves (Zenaida macroura) wintering in southern Ontario over 48 years. This used to be a rare bird, but it has apparently benefited from a warming climate, suburban habitat, and winter feeding. (b) The population of mallards (Anas platyrhynchos) wintering in southern Ontario over 35 years, illustrated with two independent sets of data. This duck has expanded its breeding and wintering ranges into eastern Canada, likely in response to habitat made available by the clearing of forest. (c) The population of lodgepole pine (Pinus contorta) near Snowshoe Lake, British Columbia, during natural afforestation following deglaciation 7000-9000 years ago. In this case, tree populations are indicated by the amount of pollen in dated layers of lake sediment. Sources: Modified from (a) Freedman and Riley (1980); (b) Goodwin et al. (1977); (c) MacDonald and Cwynar (1991).
Figure 9.2. Population Growth Stopping at Carrying Capacity. (a) The population growth of yeast cells grown in a flask is initially exponential but then levels off at the carrying capacity of the habitat. Carrying capacity is determined by the volume of the flask, the quantity of nutrients available, and the increasing concentrations of toxic metabolites, including ethyl alcohol. (b) The population of a moss colonizing a suitable, but initially bare, rock substrate in Iceland. The carrying capacity is limited by the amount of two-dimensional space. Sources: Modified from (a) Krebs (1985); (b) Silverton (1987).
Figure 9.3. Population Growth and Crash. In 1910, reindeer (Rangifer tarandus tarandus; the Eurasian subspecies of caribou) were introduced to two islands in the Aleutian chain off Alaska in an attempt to establish a new food resource for local use. On both islands, the reindeer population increased rapidly. However, they exceeded the carrying capacity of the habitat and caused severe damage through overgrazing. The populations then crashed. Source: Modified from Krebs (1985).
In Detail 9.1. Exponential Growth
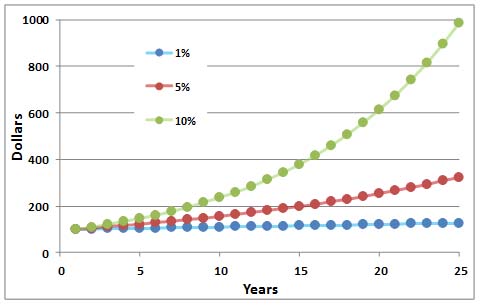
A constant rate of increase leads to rapid growth in the size of a population. This happens for the same reason that money invested at a fixed rate of interest will quickly increase in quantity. This phenomenon, known in finance as compound interest, is illustrated below.Consider, for example, an investment of $100 made at a fixed interest rate of 10% per year, locked in for a 10-year period. After Year 1, the initial deposit grows to $110, which represents the initial investment plus accumulated interest. In Year 2, the 10% interest rate is applied to the $110, so the earned interest is larger ($11) than in Year 1 ($10). In Year 3, the 10% interest is applied to the accumulated $121, so the earned interest is larger yet ($12.10), and the accrued value of the investment is $133.10. At the end of Year 4, the initial investment of $100 is worth $146.41. It is then $161.05 at the end of Year 5 … and $259.37 at the end of Year 10, representing an impressive 159% return on the initial investment. Clearly, a compounded rate of interest leads to a rapid increase in capital.
Exponential growth refers to the accelerating growth of an initial quantity due to a constant rate of increase. Sometimes an important parameter known as the doubling time is calculated—it is the time required for a two-fold increase in capital. The doubling time can be roughly calculated as 70 divided by the constant rate of increase. In the example above, 70 divided by 10% per year yields seven. Therefore, the initial $100 would double in amount in only seven years, and the accumulated $200 would again double (to $400) in another seven years, and so on as long as the investment conditions do not change.
The mathematics of compounded interest can also be applied to the exponential growth of populations of organisms. One example will suffice: In 2015, the global human population was about 7.3 billion people, growing at about 1.3% per year. Therefore, in only 54 years (i.e., 70 divided by 1.3% per year), the human population could double to more than 14 billion, if the growth rate were not to change. The environmental implications of such a population increase are immense (see Chapter 10).
Figure 9.4. Exponential Growth. This curve shows the growth of an initial deposit of $100 invested at a compound interest rate of 1, 5, or 10% per year. Biological populations also grow in an exponential fashion if their rate of increase is constant. However, when resources become limiting, the rate of increase decreases, and the population may crash.
Community Ecology
An ecological community is an aggregation of populations that occur in the same time and place as a distinctive grouping, and that interact physically, chemically, and/or behaviourally. The study of relationships among species within communities is known as synecology. Strictly speaking, a community consists of all plant, animal, and microbial populations occurring together on a site.
The Niche
Each species within a community exploits the environment and interacts with other species in a particular manner. Ecologists use the word niche to describe the role of a species in its community, which can also be viewed as its “occupation” or livelihood. Some niches are relatively narrow and specialized, as is the niche of bats that feed only on flying insects of a certain size, or wasps that pollinate only one or a few species of plants. Other niches, however, are much broader, such as those of bears and humans, both of which forage over an extremely broad range and affect their ecosystem in diverse ways.
The so-called fundamental niche is determined by the range of a species’ tolerance of environmental conditions. These tolerances are reflected in the ways that a species obtains its nutrition and how it interacts with other species, and they are mediated by aspects of behaviour, morphology, and physiology. In comparison, the realized niche reflects the range of environmental conditions that a species actually manages to exploit in nature. The realized niche is smaller than the fundamental niche because all species are to some degree constrained by biological interactions such as competition, predation, and disease.
Functional Communities
Because of their complexity, entire communities are rarely examined by ecologists. Ecological studies are usually limited by the amounts of funding and breadth of expertise available. Instead, community-level research usually involves the examination of selected groups of similar organisms, such as “communities” of insects, fish, birds, plants, or microbes. Although the scope of such work is limited, it does allow ecologists to investigate important aspects of community ecology.
Forest communities, for example, contain a wide range of organisms of various species and sizes, including plants, animals, and microorganisms. The populations of the diverse species interact in myriad ways. Trees, for instance, provide the physical structure of the habitat, make food available for herbivores, and drop leaf litter that is decomposed by species of the detrital food web. Other interactions within a forest community include predation, parasitism, and disease, as well as symbioses such as pollination, seed dispersal, and root mycorrhizae. Because of the inherent complexity of forest communities, most ecological studies only investigate selected components.
This pragmatic approach to community-level research can be illustrated by studies of the ecological effects of forestry conducted in the Maritimes by the author and a number of students. To do this work we divided the larger community into the following functional groups:
- trees, which we defined as woody plants with a diameter greater than 10 cm
- shrubs, with a diameter less than 10 cm but taller than 1 m, including shrub-sized young individuals of tree species as well as “true” shrubs
- ground vegetation, including all plants, mosses, and lichens, growing within 1 m of the ground
- epiphytes growing on other plants, such as lichens and mosses on the bark-covered surfaces of trees
- small mammals such as mice, shrews, voles, and squirrels
- large mammals such as deer, bear, and coyote
- birds
- reptiles and amphibians
- insects
- fungi and other microorganisms in the soil
During some of the studies of birds, specific work was done with species that nest in cavities in trees. These comprise a “cavity-requiring” element of the larger avian community. Similarly, work on insects and other invertebrates has involved functional groups that live in soil, in rotting deadwood, or on foliage. But even with all of these (and other) functional groups, we did not manage to examine all of the elements of the forest communities that we were studying.
Factors Influencing Communities Ecological communities are affected by various environmental factors, particularly those described below.
Species Present – Obviously, only those species that are present in a habitat, or are capable of dispersing into it, can play a role in the community that develops. The ability of a species to colonize an available habitat is influenced by its biology, intervening barriers such as a mountain range or ocean, the disturbance regime, and other factors. Increasingly, humans are influencing the species composition of communities, often by introducing non-indigenous species beyond their natural range.
Appropriate Habitat – If a habitat is unsuitable, then a particular species will not be able to use it even if it is capable of dispersing to the site. There are many aspects of habitat suitability, and all of them must be satisfied within the limits of tolerance of a species if it is to become a component of a community.
Biological Interactions – Species interact through herbivory, predation, competition, disease, and symbiosis, the latter including mutualism, commensalism, and parasitism. All of these interactions can influence the presence and abundance of species within communities. The following examples illustrate these influences.
Herbivory – occurs when animals feed on plant biomass. Larvae of the hemlock looper (Lambdina fiscellaria) are voracious feeders on the foliage of spruce, fir, and other coniferous trees. When conditions are suitable, this moth can proliferate rapidly, causing damage over a large area of forest, as periodically happens in eastern Canada. Stands defoliated for several years have many dead trees, representing an important element of community change. The loss of much of the forest canopy has indirect effects, such as allowing understorey plants to grow more vigorously. The changes in vegetation affect the habitat available for species of insects, birds, and other animals. Microorganisms and other detritivores are also affected because large quantities of dead tree biomass are available to be decomposed.
Predation – involves an animal killing and eating another animal. Predators can greatly reduce the abundance of their prey, thereby changing the structure of the community. For instance, during the summer, most forest birds feed on insects, spiders, and other invertebrates, which are nutritious food for both adults and their rapidly growing nestlings. Avian predation can change the invertebrate community, as has been demonstrated by studies in which small areas of forest were enclosed with netting. This excluded avian predators, but invertebrates could move in or out. Under these conditions, the abundance of many insects and spiders increased, with species vulnerable to avian predation benefiting the most.
Image 9.4. Species interact with each other in various ways, such as herbivory, predation, competition, disease, and symbiosis. This photo shows caribou (Rangifer tarandus) grazing in a tundra meadow in the Nunavik region of northern Quebec. Source: B. Freedman.

Competition – occurs when the biological demand for an ecological resource exceeds the supply, causing organisms to interfere with each other. Plants, for example, often compete for access to limited supplies of sunlight, water, nutrients, and space. Animals may compete for food, nesting sites, mates, and other resources. Intraspecific competition occurs when individuals of the same species vie for access to resources, while interspecific competition occurs between species. If a species is particularly effective at co-opting resources to its own benefit, it may displace other species, a phenomenon known as competitive displacement (or in extreme cases, competitive exclusion). This affects the presence and relative abundance of species in the community. For example, sugar maple (Acer saccharum) is a highly competitive tree in hardwood forests of eastern Canada. Where environmental conditions are well suited for this species, it can dominate mature stands. If large sugar maple trees are removed from a stand, perhaps by a selective timber harvest, other tree species (as well as small sugar maples) will benefit from the reduced competition and will grow more vigorously.
Disease – is a pathological relationship in which the health of plants or animals suffers from an infestation of another species, usually a microbe. Virulent diseases can cause enormous changes in the composition of ecological communities. In the early 1900s, the American chestnut (Castanea dentata) was afflicted by chestnut blight (Endothia parasitica), an introduced fungal pathogen. Because chestnuts have little immunity to this disease, the species was virtually eliminated from the forests of eastern North America by the 1950s. This change released other tree species from competition with the previously dominant chestnut, and they quickly filled gaps in the canopy created by its demise.
Symbiosis – refers to intimate relationships that may occur among species. This may involve an obligate relationship in which the symbionts cannot live apart, but more commonly the association is somewhat flexible. Symbioses can greatly influence the performance of species in particular environments by improving their competitive ability and decreasing their vulnerability to predation, disease, or other stresses.
The main types of symbiosis are mutualism, in which both partners benefit; parasitism, in which one organism benefits and the other is harmed; and commensalism, in which one organism benefits without harming the other. While symbioses are critical to one or both partners, they can also indirectly affect the habitat and the resources available to other members of the community.
Lichens are a familiar example of a mutualism. They are an obligate association between a fungus and either an alga or a blue-green bacterium. The fungus benefits from the productivity of the photosynthetic partner, while the latter gains a relatively moist microhabitat and improved access to inorganic nutrients.
Another mutualism, called a mycorrhiza, is an intimate association between fungi and the roots of vascular plants. The plant benefits through enhanced access to nutrients, especially phosphate, while the fungus receives nutritious exudates from the roots. This mutualism also provides a broad, community-level benefit through increased primary productivity. Many species of legumes live in a mutualism with the bacterium Rhizobium japonicum, which fixes nitrogen gas (N2) into ammonia, a critical nutrient.
Another mutualism involves species of dinoflagellates (single-celled algae) that live within corals (small, colonial animals), where they receive protection and access to nutrients. The corals benefit through access to the photosynthetic productivity of the algae.
Many animals eat plant biomass, but few are able, on their own, to digest complex polymeric biochemicals such as cellulose and lignin. Consequently, many herbivores live in a mutualism with microorganisms, which inhabit their gut and secrete enzymes that digest cellulose and lignin, making those abundant sources of nutrition available to the animal. Cows, deer, and sheep host their digestion-aiding microorganisms in a specialized pouch of their fore-stomach, called the rumen. Humans also harbour a diverse community of microorganisms in their gut, many of which are important to our nutrition.
Other mutualisms include the many species of flowers that are pollinated by particular kinds of insects. Pollination is crucial to the reproductive success of plants, while the insects benefit from an abundant food source of nectar or pollen. In addition, herbivores in the community benefit from the fruits that are produced because of pollination, and in turn they may help to disperse the plant seeds.
An example of commensalism is the epiphyte community of plants, lichens, and mosses that often grow on large trees. The epiphytes gain an benefit from the relationship through increased access to sunlight, but the host trees are not affected to any meaningful degree. There are many familiar examples of parasitism, including fleas on a dog and tapeworms in humans. The parasite benefits by taking nutrition from the host, but the host usually suffers, and may even die from a severe infestation.
Image 9.5. A mutualism is an intimate symbiosis in which both partners benefit from the relationship. Lichens, such as the light-coloured Parmelia saxatilis in the photo, are an obligate mutualism between a fungus and an alga, meaning the two species cannot live apart in nature. Thus, taxonomists treat them as a single “species.” Source: B. Freedman.

Disturbance is an event of destruction of some part of a community, an occurrence that is followed by a sometimes pronged period of ecological recovery called succession. All communities are dynamic, changing over time in their species composition and functional attributes (such as productivity, decomposition, and nutrient cycling). However, the rate of change depends on the stability of environmental conditions, which is greatest in communities that are close to the end-point of a succession. In contrast, the most dynamic communities are associated with the younger stages of succession. Disturbances can occur on two spatial scales.
- Stand-replacing disturbances are caused by wildfire, a disease epidemic, clear-cutting, and other cataclysmic events. This kind of disturbance is extensive and results in the immediate replacement of a community with a different one, followed by a period of successional recovery. Over time, succession may regenerate a community similar to what existed before the disruption, or a different one may result. The younger stages of a sere (successional sequence) are especially dynamic in terms of community change. During the initial years of recovery, competition is not intense, and ruderal, r-selected species dominate. Later stages of succession are much less dynamic, and K-selected species dominate.
- Microdisturbances are local disruptions that affect small areas within an otherwise intact community. A microdisturbance may, for instance, be associated with the death of an individual large tree, which results in a gap in the canopy, below which community change is relatively dynamic as species compete to take advantage of the additional sunlight. Similarly, the death of an individual coral head represents a microdisturbance within a tropical reef community. Although ecological changes are dynamic within a gap created by a recent microdisturbance, at the stand level the community is relatively stable. Gap-phase community dynamics occur in all ecosystems but are especially important during later stages of succession, such as in older-growth forests.
Image 9.6. Ecosystems are occasionally subjected to catastrophic disturbances, such as these forest fires in 2002 in the boreal forest of northern Quebec. The individual fires are marked with a red dot, and their smoke plumes are blowing to the south. The large white mass at the bottom right is cloud cover. Source: NASA photo ID 751339; https://www.dvidshub.net/image/751339/fires-quebec-canada-send-smoke-us-natural-hazards#.VOS_vXUtHIU

Spatial variation of the environment reflects the fact that conditions are always changing from place to place, and sometimes extremely so. These spatial variations influence the character of ecological communities, in ways that may be gradual or more rapid:
- Gradual changes environmental conditions are associated with varying altitude on a mountain, differences of climate over large distances across continents, and other relatively continuous gradients. This type of spatial change is reflected in gradual variations of communities because individual species have different but overlapping tolerances and requirements of environmental conditions. These biological differences result in overlaps of the distributions of species, which can make it difficult for ecologists to determine the locations of boundaries (ecotones) between types of communities.
- Rapid changes in environmental conditions occur at sharp boundaries between different kinds of soil or bedrock, at interfaces between aquatic and terrestrial habitats, and in places affected by disturbance. The latter influence can occur, for instance, between a burned and unburned tract of forest, or between an ecological reserve and its surrounding area, which may be affected by agriculture or forestry. Relatively discrete changes in environmental conditions favour large differences in community types, with distinct boundaries between them.
Landscape Ecology
Landscapes (or seascapes in the marine context) are a mosaic of “patches”, each of which represents an ecological community. A landscape may contain various kinds of communities for the following reasons:
- each community reflects particular environmental conditions, such as different soil or bedrock types or variations of standing water (as in lakes, streams, or wetlands)
- the communities represent various stages in succession, such as patches of different age after wildfire or insect damage
- the communities may be related to land-use, as when parts of landscapes are affected by urbanization, agriculture, forestry, roads, or other human influences.
Over time, the spatial patterns of communities on landscapes are highly dynamic. This largely reflects the influence of disturbances and successional recovery. A patch that today is a pasture, a recent clear-cut, or a burn may be a mature forest after 50 years of succession. Similarly, a pond may in-fill over the centuries and become a wetland, which with further time may succeed into a forest. Ecologists use the term “shifting mosaic” to integrate the spatial and temporal variations of communities on landscapes. The following factors affect the shifting mosaic of communities.
- Patch size relates to the area of particular stands of communities (a stand is a community in a specific place). All species need some minimal area of habitat to support their populations, and small patches may not be adequate for that purpose. Relatively small patches may, however, help to support a population living in several stands on the landscape (an extensive population of this sort is known as a metapopulation). This can happen if the patches are connected by corridors to other suitable habitat, or if the species is capable of dispersing through surrounding inhospitable habitat (for this to occur, the landscape matrix must be permeable to movements of the species).
- The amount of edge is important because it influences the length of ecotone (transitional) habitat associated with a patch. A circular patch has the smallest ratio of edge to area, and smaller patches have higher ratios than larger ones of the same shape. An ecotone between patch types is a particular kind of habitat, and it may be selectively used by “edge species.” However, the greater the ratio of edge to area, the less “interior” habitat there is (this is uninfluenced by ecological conditions associated with an ecotone). Ecologists have identified “interior species” that are less successful if they try to use habitat close to an edge. Certain forest birds, for example, experience greater rates of predation and nest parasitism in small remnants of mature forest (see Chapter 26).
- Connectedness refers to the presence of links between otherwise discrete patches of similar habitat. These links may be used by a species as corridors to move among patches, allowing their metapopulation to function on the landscape. As was noted previously, connectedness is also related to the ability of a species to disperse among habitable patches through the surrounding habitat.
- Age-class adjacency is important in a landscape in which patches represent different stages of a successional sequence. This commonly occurs in landscapes affected by disturbances such as wildfire, insect epidemics, or clear-cutting. In general, patches of a similar post-disturbance age will be comparable in many aspects of habitat, while those of different age will be less similar. This can be an important consideration for movements of species among isolated patches that are suitable as habitat.
- Complex habitat requirements are characteristic of some larger animals, such as deer, bear, and wolf. These species need different kinds of habitat patches for specific purposes at various times of year. Because these animals participate in various kinds of communities, all of the habitat patches they need must be present on the landscape if a viable metapopulation is to be sustained.
- Landscape-level biodiversity is related to the richness of community types over a large area (see Chapter 7). A landscape that is uniformly covered by a single community has less biodiversity at this scale than one composed of a rich and dynamic mosaic of different kinds of communities.
- Landscape-level functions operate over extensive areas, and they may integrate the influences of many kinds of communities. A watershed, for example, is the expanse of terrain from which water drains into a stream, lake, or some other waterbody. Most watersheds contain various kinds of habitat patches, each with particular influences on hydrology and water chemistry. In general, watersheds covered with mature forest yield the cleanest flows of water. Other environmental services provided by well-vegetated landscapes include evapotranspiration, control of erosion, moderation of climatic extremes, and absorption of atmospheric carbon dioxide and release of oxygen.
Landscape ecology is an important subject area in environmental science. Humans commonly affect individual stands of particular kinds of communities, but many of the ecological effects must be managed at the scale of landscapes and seascapes.
Image 9.7. A landscape is a mosaic of various kinds of communities, each stand of which represent a patch. In addition, landscapes are subjected to patch dynamics associated with natural disturbances, such as wildfire, windstorms, and insect outbreaks. However, the patch dynamics of many forested landscapes are being increasingly structured by forestry. In this aerial view of an area in New Brunswick, the natural forest is being harvested by clear-cutting (the lighter patches are snow in clear-cuts), which initiates a succession that restores a forest for another harvest in 60-80 years. Unless some areas are set aside for protection, this entire landscape may become used in this way. Source: M. Sullivan.

The Biosphere
The biosphere consists of all life and ecosystems on Earth. It is bounded by the presence of living organisms, and it is the only place in the universe definitely known to support life. Ecological processes at the level of the biosphere include global climatic, oceanic, and atmospheric regimes (Chapter 3), the planetary energy budget (Chapter 4), and global nutrient cycles (Chapter 5). These biospheric processes influence all life and ecosystems. At the same time, life and ecosystems also influence biosphere-level processes.
In fact, some scientists have suggested that there may be a degree of homeostatic control, or feedback, between the reciprocal influences of global ecosystems and their environment. A notion describing these biosphere-environment relationships is known as the Gaia hypothesis, a controversial idea popularized by the scientist James Lovelock. He suggests that organisms and ecosystems have caused large changes to occur in certain physical and chemical attributes of the global environment, and they have resulted in improvements in living conditions on the planet. The hypothesis envisions all of Earth’s species and ecosystems as being a sort of “superorganism,” which is called Gaia. According to Lovelock, Gaia attempts to optimize environmental conditions toward enhancing its own health and continuity, and it uses feedback mechanisms to help maintain conditions within a range that life can tolerate. The ancient Greeks believed that Gaia (or Gaea) was the prolific ancestor of many of their most important gods. The Romans, who adopted many Greek gods and ideas, knew Gaia as Terra. More recently, the Gaian myth has been personified as “mother Earth.”
The Gaian idea is attractive and interesting, largely because it integrates many ideas and large-scale observations into a consolidated belief and world view. However, Earth is the only planet in the universe that is known to support life and ecosystems, and so it is the only known replicate in the great experiment of life. Consequently, the Gaia hypothesis cannot be tested by rigorous experimentation, and for this reason many scientists reject its inferences. Except in the broadest of terms, Gaian ideas may not be useful in helping humans to manage the detrimental impacts of their increasing population and industrial activities on the biosphere. Nevertheless, some intriguing lines of evidence can be marshalled in support of the Gaian notion. Two examples follow.
Atmospheric Oxygen
Earth’s primordial atmosphere did not contain oxygen (O2). This gas appeared only after the first photosynthetic organisms, blue-green bacteria, evolved. These, and the somewhat later evolved green algae, give off O2 as a waste product of photosynthesis. The modern concentration of O2 in the atmosphere, about 21%, has resulted entirely from photosynthesis and is a critical environmental factor for most species and many key ecological processes. It appears that the concentration of atmospheric O2has been fairly stable for several billions of years. This suggests a long-term equilibrium between O2production by photoautotrophs and its consumption by respiration, including decomposition. Interestingly, if the concentration of oxygen were much higher than 21%, say 25%, then biomass would be much more combustible. This condition would lead to more frequent and extensive wildfires, which would severely damage terrestrial ecosystems. These observations can be interpreted as suggesting the existence of a homeostatic control of the concentration of atmospheric O2, operating at the biospheric scale. This control may achieve a balance between the need to have sufficient O2 to sustain the most abundant organisms (which have an aerobic metabolism), and larger O2 concentrations that would result in destructive conflagrations.
The Greenhouse Effect
The concentration of carbon dioxide (CO2) in the atmosphere is regulated by a complex of physical and biological processes by which this gas is emitted and absorbed. Atmospheric CO2 is important in Earth’s greenhouse effect, which maintains the surface temperature within a range that organisms can tolerate (Chapters 4 and 17). The greenhouse effect helps maintain an average surface temperature of about 15°C, compared with the -18°C that would otherwise occur and would be too cold for organisms to tolerate. Advocates of the Gaia hypothesis suggest that these observations imply a homeostatic control of atmospheric CO2 and an indirect control of the greenhouse effect and climate.
There is clear evidence that organisms and ecosystems cause substantial changes to occur in their environment, and also that they are affected by those conditions. The scientific community does not, however, widely support the notion that Earth’s species and ecosystems have somehow integrated into a mutually benevolent symbiosis aimed at maintaining a comfortable range of environmental conditions.
The Gaia hypothesis is nevertheless quite useful in environmental science. Gaian ideas emphasize the diverse connections that exist within and among ecosystems, as well as the damaging consequences of human actions that are increasingly causing large environmental and ecological changes to occur. If these changes were to exceed the biospheric limits of homeostatic tolerance and repair, the consequences for the planet’s geophysiology and ecology could be catastrophic.
Evolutionary Ecology
Evolutionary ecology is a fusion of ecology and evolution – it involves the interpretation of ecological relationships in terms of evolution, natural selection, and related themes. Moreover, it acknowledges that species and their ecology have an evolutionary history that involves change over time, usually occurring as adaptive responses to environmental influences. Natural selection is a particularly significant influence on evolutionary change, and it is evident in the regimes of environmental stressors and disturbances that affect the survival and reproductive success of organisms. Core subject areas in evolutionary ecology are the evolution of life-history traits, of relations between species (such as mutualism, parasitism, pollination, and predator and prey), and of communities and biodiversity in general. Examples of the interpretation of nature in terms of evolutionary ecology include convergence and coevolution:
- Convergence occurs when unrelated species occupy similar niches in distant but comparable environments. As a result they are subjected to parallel regimes of natural selection, and evolve to be similar in morphology and behaviour. An example noted earlier is the placental wolf (Canis lupus) of Eurasia and North America and the thylacine or marsupial wolf (Thylacinus cynocephalus) of Tasmania.
- Coevolution occurs when species interact in ways that affect the survival or reproductive success, so that they are subject to regimes of natural selection that result in integrated evolutionary change. The intensity of coevolution can, however, vary greatly. In extreme cases, it may lead to the evolution of an obligate mutualism, meaning that neither partner can survive without the other. Lichens are one example—these are mutualisms between a fungus and an alga, neither of which can survive on its own.
Coevolution also affects the feeding relationships of organisms. For example, milkweed plants (such as the common milkweed, Asclepias syriaca) have evolved to produce high concentrations of cardiac glycosides, which are distasteful and poisonous to most animals and help to protect the plants from being eaten. However, the monarch butterfly (Danaus plexippus) has evolved a tolerance to those biochemicals and so can eat milkweed tissues (in fact, their larvae eat nothing else). When this happens, the cardiac glycosides are incorporated into tissues of the monarchs, giving a measure of protection from being eaten by birds and other predators. The defence is so effective that the unrelated viceroy butterfly (Limenitis archippus) has evolved a colouration and flight pattern that mimic those of the monarch, which helps deter its own predators.
Coevolutionary interactions have also been studied between many plants and their pollinating insects. One example is the bee orchid (Ophrys apifera), whose flower structure has evolved to resemble the abdomen shape of the solitary bee Eucera. This induces male bees to try to copulate with the orchid, which becomes pollinated in the process (although it is also capable of self-pollination).
Image 9.8. Convergent evolution can be illustrated by comparison of the skulls of two unrelated mammalian predators – the timber wolf (Canis lupus), a placental mammal, and the thylacine or Tasmanian wolf (Thylacinus cynocephalus), a marsupial. Source: B. Fenton.

Some ecologists have even speculated about the possibility of natural selection operating at levels higher than populations or species (this is known as group selection). For instance, there has been speculation that certain boreal-forest assemblages have evolved to be highly flammable, and that this characteristic has promoted stand-replacing wildfires, a disturbance that can rejuvenate the ecosystem. It is true that mature boreal forest dominated by black spruce or white birch can easily ignite and burn rapidly, but there is not yet a supporting theory of the adaptive evolution of communities, as opposed to their constituent species.
As is true for all of biology, evolutionary theory provides a key element of the conceptual background of ecology. Knowledge and insights from evolution and ecology are also central to understanding the environmental effects of the human economy.
Applied Ecology
The application of ecological principles to dealing with economic and environmental problems is known as applied ecology (or as environmental ecology). There are three major subject areas:
- the management of renewable resources, such as those important in agriculture, fisheries, and forestry
- the prevention or repair of ecological damages, such as those related to endangered biodiversity, and the restoration of degraded land or water
- the management of ecological processes, such as productivity, carbon storage, nutrient cycling, hydrology, and erosion
Later chapters in this textbook contain many examples of the use of applied ecology to deal with problems of resource management, pollution, and disturbance. Although the subject is not dealt with here in detail, it can be briefly illustrated by the following examples:
- Setting Harvest Limits: The rate of forest productivity in a region can be estimated by measuring trees in plots that are re-sampled over the years, or by coring populations of trees and examining the annual ring-width of the recently grown wood. This information can be used to set limits on the amount of timber that can be harvested without degrading the resource. Analogous methods of measuring productivity can be used to manage the sustainable harvesting of populations of mammals, birds, and fish.
- Increasing Biological Productivity: Applied ecological research can determine whether the productivity of biological resources can be increased—for example, if forest growth could be enhanced by applying fertilizer, thinning dense stands, or establishing plantations. Research can also predict other implications of these management practices, including effects on biodiversity and water quality.
- Remediation, Reclamation, and Restoration: Ecologists can provide research-based advice for improving conditions in areas that have been degraded by pollution or disturbance. Various kinds of schemes can be used to deal with ecological damage:
- Remediation involves actions that are undertaken to deal with a particular problem, such as liming lakes and rivers to decrease their acidity, planting tolerant plants in polluted environments, or undertaking captive breeding and release to increase the abundance of an endangered species.
- Reclamation involves more comprehensive actions to establish a productive ecosystem on degraded land. For example, an old landfill or a disused industrial site may be reclaimed to a permanent cover of vegetation, such as a pasture.
- Restoration has a loftier goal of attempting to establish a self-maintaining facsimile of a natural ecosystem on degraded land. For instance, the techniques of restoration ecology might be used to convert abandoned farmland back to a native prairie or forest.
- Offsets for Greenhouse Gases: Ecologists can predict the amount of forest that must be grown and protected in order to offset industrial emissions of greenhouse gases. This can allow companies and nations to reduce their net emissions of CO2 and to make progress toward meeting international agreements to deal with climate change.
- Reducing Erosion: Some areas have been badly degraded by erosion caused by deforestation and other land-use changes. Ecologists can find ways to deal with this problem—for example, by planting forests, using terraced agricultural fields, or managing local hydrology to reduce overland or subsurface water flows.
Conclusions
Ecology is the study of the relationships of organisms with the environmental factors that provide the requirements of life and livelihood. These factors include resources such as nutrients and food, the influences of other organisms through competition and predation, as well as stressors such as disturbance and pollution. Knowledge of ecology is central to understanding many of the most important damages that the human economy is wreaking to the biosphere. Applied ecological knowledge is essential to managing renewable resources on a sustainable basis, to conserving biodiversity, and to avoiding and repairing damage caused by pollution and other destructive influences of humans.
Questions for Review
- Distinguish between autecology and synecology. Give examples to illustrate each.
- What are birth rate, death rate, immigration rate, and emigration rate? How do these demographic factors influence changes in a population?
- What environmental and biological influences affect the structure and function of a kind of ecological community with which you are familiar?
- What are the attributes of an ecological landscape (or seascape)? Illustrate your answer with an example in the region where you live.
- In a general sense, what are the goals and methods of a project in restoration ecology?
Questions for Discussion
- Use the principles of autecology to discuss the resource needs and environmental tolerances of humans.
- How is the human economy integrated into the biosphere?
- How are ideas and knowledge about population ecology relevant to people?
- What are the core elements of the Gaia hypothesis? What evidence exists to support this hypothesis? How are Gaian ideas relevant to environmental science?
- It is difficult and expensive to implement a large-scale project in restoration ecology. Under what sorts of conditions do you think such a project might be worthwhile?
Exploring Issues
- The niche has sometimes been described as the “occupation” of a species—what it does for a living, the resources it uses, and its habitat. You have been asked to make a presentation to a group of non-ecologists, in which you must describe the niche of humans. What information would you include in your presentation?
References Cited and Further Reading
Begon, M., R.W. Howorth, and C.R. Townsend. 2014. Essentials of Ecology. 4th ed. Wiley, Cambridge, UK.
Freedman, B. and J. Riley. 1980. Population trends of various species of birds wintering in southern Ontario. Ontario Field Biologist, 34: 49-79.
Freedman, B., J. Hutchings, D. Gwynne, J. Smol, R. Suffling, R. Turkington, R. Walker, and D. Bazeley. 2014. Ecology: A Canadian Context. 2nd ed. Nelson Canada, Toronto, ON.
Goodwin, C. E., B. Freedman, and S. M. McKay. 1977. Population trends in waterfowl wintering in the Toronto region, 1929–1976. Ontario Field Biologist, 31: 1-28.
Grime, J.P. 2001. Plant Strategies, Vegetation Processes and Ecosystem Properties. 2nd ed. J. Wiley, London.
Harris, J.A., P. Birch, and J. Palmer. 1996. Land Restoration and Reclamation: Principles and Practices. Addison-Wesley, Reading, MA.
Kimmins, J.P. 2003. Forest Ecology. 3rd ed. Prentice Hall, New York, NY.
Krebs, C.J. 2008. Ecology: The Experimental Analysis of Distribution and Abundance. 6th ed. Benjamin Cummings, New York, NY.
Lovelock, J.E. 2000. Gaia: A New Look at Life on Earth. Oxford University Press, New York, NY.
MacDonald, G.M. and L.C. Cwynar. 1991. Post-glacial population growth rates of Pinus contorta ssp. latifolia in western Canada. Journal of Ecology, 79: 417-429.
Mills, S. 1996. In Service of the Wild: Restoring and Reinhabiting Damaged Land. Beacon Press, Boston, MA.
Odum, E.P. 1997. Ecology: A Bridge Between Science and Society. Sinauer Associates, New York, NY.
Odum, E.P. and G.W. Barrett. 2004. Fundamentals of Ecology. Brooks Cole, Florence, KY.
Silverton, J. and D. Charlesworth. 2001. Introduction to Plant Population Biology. Blackwell Science Ltd, Oxford, UK.
Smith, L.E. 1991. Gaia: The Growth of an Idea. St. Martin’s, New York.
